A Triumph with a Triumphans
Successful seed set in cultivated Philodendron triumphans/sp. nov. ”Chocó Red”
by Jay Vannini

Detail of an upper petiole section and lower leaf lamina of a mature Philodendron sp. nov. “Chocó Red”. Author’s plant and image.
OK, so it’s only a minor triumph.
Despite their widespread and long-standing popularity in cultivation, production of hemiepiphytic Philodendron species from artificially propagated seed remains a surprisingly uncommon event. Likely due to irregular or seasonal flowering cycles (Croat, 1997), evidence of self-incompatibility in some species (Maia et al., 2010), and the comparative difficulty in successfully pollinating inflorescences with refrigerated pollen subjected to prolonged storage (pers. obs.), significant challenges exist to dependable seed production of these plants in northern countries. Asexual propagation–either from green stem sections or via plant tissue culture (PTC)–remains the preferred method that collectors and commercial nurseries use to greatly increase their numbers of marketable scandent philodendrons.
One noteworthy exception involving a localized rarity was that of Philodendron patriciae, which produced a large seed crop from hand-pollinated wild origin founders in 2012 at the Huntington Botanical Garden in southern California. Dylan Hannon, a curator there who produced this seed, published on this event following seed and seedling distribution to other collections around the world (Hannon, 2014). A follow-up cross of an old, wild-collected P. patriciae seed parent and a Huntington F1 pollen donor was made by me in late 2018, and later still into full F2 by Chris Hall and Arden Dearden of Queensland, Australia using artificially propagated, seed grown stock for both parents. Other more common but still desirable Philodendron species in ornamental horticulture are also produced from seed in Australia on a fairly regular basis (A. Dearden, pers. comm.). Simple and complex Philodendron climbing hybrids have surfaced sporadically around the world since the mid-20th century (Graf, 1980) and some of the better of these continue to attract the interest of both commercial nurseries and collectors.

Mature Philodendron spiritus sancti flourishing in a research greenhouse at a public garden in the U.S.. Author’s image.
Another highly coveted species in horticulture that is reportedly subject to ongoing wild collection pressure is Philodendron spiritus sancti. Plants propagated from stem cuttings regularly fetch five figures (USD/EUR) on global markets. Reports of seed production in cultivated P. spiritus sancti in Brazil in 2019/2020 have been followed by other still unsubstantiated claims of artificially propagated seed and seedlings produced by plants of this species held in the U.S. and Asia. Because images of verified young seed-grown P. spiritus sancti are elusive, it remains to be seen whether the seed and seedlings being sold at this time develop true to form or are revealed as either lookalike taxa or manmade hybrids as they mature. If this new material is indeed genuine and the parent plants identified, it is certainly a welcome event for ex-situ rare aroid conservation.
Unlike other aroid genera that flower frequently and are easily pollinated such as Anthurium, Spathiphyllum, Aglaonema and Amorphophallus (Henny et al., 2004), many hemipepiphytic and vining epipetric philodendrons present timing and other logistical challenges to would-be home and greenhouse propagators (Croat, 1997; pers. obs.). Information available from published sources and online generally emphasizes success with woody-stem, landscape suitable species that extrude pollen in dry strands, now segregated into the genus Thaumatophyllum (Griffith Jr., 2001; Miller & Newton, 2006; Sakuragui et al., 2018). Plants from this genus, both species and hybrids, are relatively easy to produce from seed. Philodendrons with resinous pollen are notably more difficult to work with unless certain techniques are employed. Detailed information on Philodendron reproductive biology is mainly available from field studies (e.g., Maia et al., 2010), so any information on the flowering phenology of lesser-known species that private collectors and botanical gardens can provide to aroid researchers is of definite value.
Despite the hurdles mentioned above, and as is the case for other aroid genera and from a plant conservation and sustainability perspective, by now country of origin nurseries should be producing the majority of their “rare” philodendrons destined for export from seed–or divisions of seed-grown plants produced in-house–rather than relying on continual harvest of wild-collected stems and whole plants.

A young Philodendron sp. nov. “Chocó Red” showing it attractive subvelvety upper leaf surfaces and (still) colorful lowers. Author’s plant and images.
A medium sized Philodendron species considered to be very desirable among specialty plant collectors is currently known under the trade names P. sp. “Chocó Red” and P. sp. “#5”. Despite key morphological differences, they are still confused with P. verrucosum by some houseplant sellers. This is very likely a plant first introduced to cultivation during the late 19th century as Philodendron triumphans (Gardener’s Chronicle, 1898; Vannini, 2020). Its most popular commercial name is a bit of a misnomer, since the showy red lower leaf surfaces that are its nicest feature when small fade rapidly to gray green in late adolescence and beyond. Based on whole leaf characters this species has been placed in Philodendron section Philodendron, subsection Achyropodium, which includes other well-known ornamental species with scaly petioles such as P. verrucosum, P. serpens, P. squamicaule and P. fibrosum. Despite a lack of reliable provenance but as the commercial name implies, it is believed to be native to lower or middle elevations of western Colombia. Regional nurseries who export it suggest that it is uncommon in the wild, but no reliable information is available as to its precise origin nor conservation status in nature. As of posting date for the present article, the species has been formally described from material in U.S. horticulture by Croat & Kaufmann (ms. in press), but remains unpublished. My article will be edited following publication of that paper to reflect general adoption of their binomial, despite some vendors using it in in interim, apparently to show off their being plugged-in.


Shown above, the dramatic lower leaf color transition in Philodendron sp. nov. “Chocó Red” from juvenile to pre-flowering size. The lead image in the article shows the concolorous lower leaves on mature specimens. Author’s plants and images.
This plant has been in cultivation in the U.S. under the “Chocó Red” moniker since the very late 2000s/early 2010s and trade until very recently appears to have relied on a constant flow of wild-collected stems sold into the northern countries. Until recently, flowering in captivity seems to be a rare event. Despite it having been trafficked by imported tropical plant flippers for about a decade and being a current favorite of the collector philodendron crowd, I am unaware of any previous reports of production of domestically produced, seed grown material of this taxon.* There are now many individuals across the world who produce this fairly hardy Philodendron on a regular basis from stem cuts. Since 2019 these domestic propagates are making it to market in ever-increasing numbers. A cursory, contemporaneous (mid October 2021) review of online nurseries, tropical plant auction sites and Etsy reveals an ample supply of cutting-grown stock being offered at a variety of sizes and price points.
I have been growing this species since early 2018 from small, weathered and likely wild-origin stems sold at local orchid shows by two foreign nurseries. I have a total of five individual founders and seven specimen plants in total, with another stem-propagated specimen donated to a major U.S. botanical garden earlier this year. Based on minor variations in stem, leaf and spathe colors when grown under identical conditions, as well as cross-pollination compatibility, I am confident that at least four of the five founders represent separate clones.
*But I’m sure both social media playpens and my email Inbox will experience rapid meltdowns if this statement is proven incorrect.

Matte gray green upper leaf surface on an emergent 6.5’/2.0 m tall Philodendron sp. nov. “Chocó Red” growing in a commercial greenhouse facility at ca. 800 fc/8,600 lux exposure. Despite their ho-hum aspect when mature, juvenile plants of this species are very showy and desirable climbing aroids that are popular with collectors. Author’s plant and image.
This species is generally easy to grow. Cultivation is very straightforward under warm tropical conditions. All of my plants are grown in large vented tubs filled with a conifer bark-based substrate (no peat or coir) and are mounted upright on treefern totems. Background temperatures throughout the year range from ca. 65 F/18.3 to 95 F/35 C, with night-time relative humidity near saturation throughout the warmer months in northern California. If allowed to sprawl, as is the case with most hemiepiphytic and epipetric philodendrons, the plants will grow horizontally at a much slower pace. While I have not found this species susceptible to spider mite infestations in a humid greenhouse, plants held in my home for propagation purposes had their leaves brown tipped and their leaf surfaces lightly damaged by spider mites in less than a month, so this should be taken into account for plants destined to spend their lives in dry interiors.
Lower (abaxial) leaf surface on a Peruvian specimen of Philodendron verrucosum that mimics the ontogenetic red-to-green leaf color transformation seen in P. sp. nov. “Chocó Red”. Immature lower leaf surfaces on this plant are also dark red. Author’s plant and image.
Despite the fact that they do not appear especially closely related, as is the case with many local types of Philodendron verrucosum (especially in the southern part of its range - see right), the leaves of P. sp. “Chocó Red” undergo a significant change in color as they mature. Unlike P. verrucosum the upper (adaxial) leaf surfaces also change texture with maturity. Small offsets begin life with vivid red lower (abaxial) leaf surfaces and subvelvety upper leaves. As leaves increase in size, they begin to lose this attractive red suffusion. By maturity both leaf surfaces are matte-subglossy, somewhat thickened in texture, and gray green in color. Fresh cataphylls and prophylls in my plants retain the bright red of young leaves and add a nice touch of color to otherwise green mature plants. Inflorescences are pure white on early emergence. Fully developed inflorescences are large (to 10”/25 cm overall), showy when open, and emerge in staggered groups of one to three from individual upper leaf axils. The peduncle is short (2.25-4”/6- 10 cm in length) and vividly striped in pink and white onto the spathe limb. Outer spathe color is pale green with a showy pink or bright red margin when furled. The lower spathe color over the floral chamber changes from green to being suffused with dark pink several weeks ahead of flowering, and suffuses deeper red a day or so prior to unfurling. Inner spathe color on opening was almost entirely bright red throughout in all my plants by the end of the first day but is reported to be magenta and white in the single inflorescence mentioned in the formal description, so it can be variable (Croat, pers. comm.). Pure white adhesive pollen is extruded over a brief period (<one hour) in short curls; sequentially from base to apex. Fresh pollen changes color and texture to a resinous amber color within 30 minutes exposure to air.


Shown above left, newly-opened inflorescence on Philodendron sp. nov. “Chocó Red”. The bright red inner spathe color intensifies over the course of the following 24 hrs to overtake the pale spathe tip. Right, newly-opened inflorescence on a Central American P. verrucosum. Note the differences in outer spathe surface textures.
Based on observations of multiple inflorescences on four flowering plants in my collection during 2021, northern hemisphere flowering begins in late spring (May), peaked from mid-June through early July, continued sporadically through until mid-August, then commenced again in the fall (October). One to three inflorescences per leaf axil were produced by my plants and opened with a ca. 10-day lag between individual inflorescences. Spathes are large, colorful and–again–unlike Philodendron verrucosum, lack visible scales or “hairs” on the peduncle and spathe exterior.


Above left, emerging white inflorescence on a Philodendron sp. nov. “Chocó Red” in early fall 2021 showing the persistent cataphylls. Right, a quartet of developing inflorescences (one partially hidden in back) on the same plant photographed in late spring 2021. Note the very smooth upper peduncle/limb on the oldest inflorescence and compare with that on the P. verrucosum shown in a previous photograph. Author’s plant and images.
Observations of early emergent inflorescences that opened in late spring revealed that both receptivity and pollen production on spadices were strictly nocturnal events in this species. Because of this and to comfortably monitor the entire flowering cycle, a trio of plants were transferred from the commercial greenhouse where they are grown to my home in late June, 2021.
Background temperatures in a large open space in my home during the period the plants were held there for pollination ranged from night-time lows of 68 F/20 C to daytime highs of 85 F/29.4 C. Room temperatures recorded while monitored when spathes were open during darkness ranged from 69 F/20.6 C to 76 F/24.4 C.

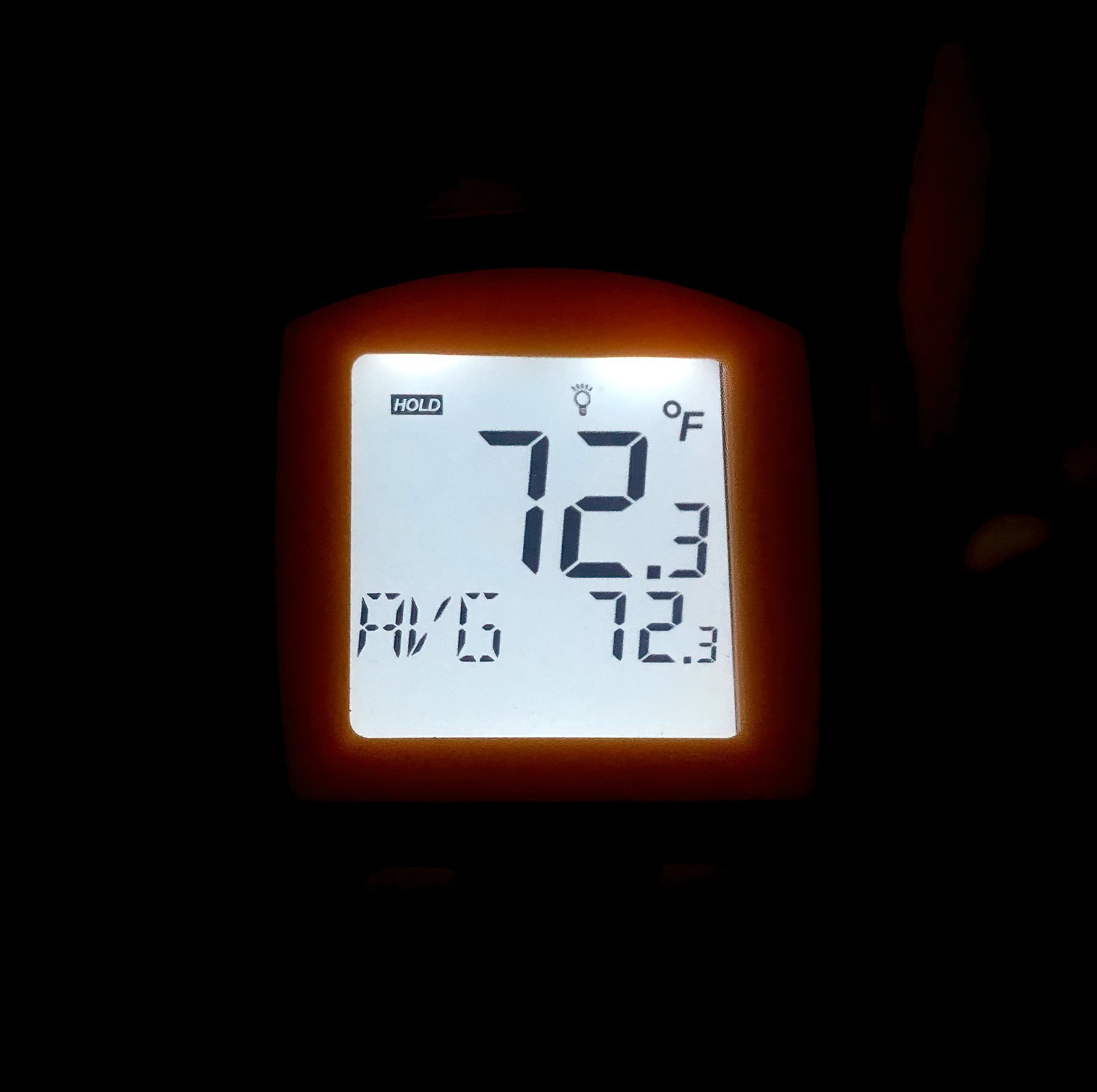
Shown above left, the industrial digital infrared thermometer employed for taking precise temperature readings of both the background environment and specific zones on an exposed spadix. Note: Fingertips are infamously bad heat sensors! Use a reliable instrument for detection of temperature changes instead. Right, room temperature shown in its illuminated digital display just after midnight prior to taking a reading off a very fragrant spadix in staminate phase just prior to pollen shed. Author’s images.
Receptivity was signaled by detectable scent emissions usually tied to increases in spadix temperature–as measured with a laser-sighted infrared industrial thermometer/heat gun–when compared to background environmental temperatures.
Scent was definitely evident at short distances when female flowers were receptive and spadix temperatures slightly higher than that of the background, but was extremely fragrant and pervasive at staminate anthesis even when the room itself was comparatively warm. An admittedly imperfect description of the spadix odor would be: “Pleasantly spicy; pungent and very peppery with sweet overtones”. Floral thermogenicity almost certainly aided in scent dispersal with peak scent evident even 15’/4.60 m away when the spadix tip recorded temperatures >10 F over background environment (e.g., an 82 F/27.8 C spadix temp vs a 72 F/22.2 C room temperature). Spadix temperatures fluctuated cyclically throughout the overnight period as was recorded during field observations of thermogenicity in Philodendron squamiferum made by Gibernau (2003) in French Guiana and observations made with several other Philodendron species by Arden Dearden in his Queensland, Australia shadehouses and gardens. A series of measurements that I made with the laser-sighted infrared thermometer over a three-week period indicated that highest temperatures were invariably located at the distal 1”/2.5 cm portion of the spadix on the second evening that the inflorescence was open for business. Temperatures of receptive pistillate sections were usually very close to those of the background environment.


Shown above left, colorful striped spathe limb on a developing inflorescence of Philodendron sp. nov. “Chocó Red”. Right, spathe cutaway to expose the receptive pistillate zone (lower) and cream-colored sterile section (above) on a spadix on the same species. Author’s plants and images.
Following removal of the upper sections of their spathes with sterile #11 scalpels, a total of four spadices on three plants were pollinated with fresh refrigerated pollen taken from other individuals between 25.06.21 and 07.07.21. The distal half of spadix was removed on all pollinated plants to minimize risk of secondary rot on the infructescence. Protective cover and increased humidity around the exposed spadix were provided by cut-down disposable clear water bottles as shown below right.
Despite apparent early success with all, damage suffered from jostling the plants on the return trip to the greenhouse caused three newly-pollinated inflorescences to fail immediately afterwards. Fortunately, the spadix pollinated last and on the smallest individual plant developed normally and the infructescence went full term to produce viable seeds harvested 101 days post-pollination.


Shown above left, a receptive spadix on a mature Philodendron sp. nov. Chocó Red” photographed in an interior of my home in the early evening hours. Right, fresh pollen extruded from the fertile staminate portion of a spadix in early morning hours. Note the very rapid color and texture change between pollen curls at the base of the staminate zone and that at the apex when exposed to air. An older pollinated spadix is shown lower left, shielded by an inverted, cutaway plastic water bottle. Note that in both these images the floral/basal chamber section of the spathe has been left intact. Spathe and spadix “tilt” on open inflorescences is commonly observed in philodendrons and is likely an adaptation to shield the flowers and pollen from rain. Author’s plants and images.
Timeline for the process noted below with times rounded to nearest half hour. Accompanying photographs document development of the infructescence.
20.06.21 - Two young mature plants moved from greenhouse to home.
22.06.21 - Plant I opened first inflorescence out of four at 0300 hrs – photographed
22.06.21 - Plant I, scent evident at 1900 hrs. Background temp ca. 70 F/21.1 C, lower spadix temperature same.
24.06.21 – Plant I, scent evident, pollen emergence at 0200 hrs and collected at same time; stored in refrigeration.
25.06.21 – Plant II opened first inflorescence at 1700 hrs; scent evident and pistillate zone on spadix glistening and receptive starting at at 1900 hrs. Background temperature 76 F/24.4 C, lower spadix temperature same. Brushed pre-treated resinous pollen on pistillate section of spadix of Plant II at 2200 hrs; then 0030 hrs and 0230 hrs on 26.06.21. Spadix covered with 1.05 pint/500 ml cutaway plastic water bottle.
26.06.21 – Third young mature plant moved from greenhouse to home for observation.
27.06.21 – Plant II, strong scent evident with pollen extruded at 0130 hrs. Background temperature 72.3 F/22.4, spadix tip temperature 92.1 F/ 32.8 C.
03.07.21 – Plant I, inflorescence II, scent evident, pistillate zone receptive starting at 1930 hrs. Refrigerated and pre-treated pollen from Plant II applied.
04.07.21 – Plant II, inflorescence II spathe opens 1630 hrs.
04.07.21-05.10.21 - Plant I, inflorescence II, scent evident and pollen extruded 2100 hrs and collected at 0530 hrs on 05.07.21; stored in refrigeration. Spadix temp at peak scent emission (two cycles separated by several hours) 94.7 F/34.8 with a room temperature of 72.3 F/22.4 C.
05.07.21 - Plant II, inflorescence II, scent evident 2000 hrs, brushed with pre-treated refrigerated pollen from Plant I that was harvested earlier the same day.
O6.07.21 – Plant III, opened first inflorescence at 1700 hrs, scent evident and pistillate zone glistening and receptive. Brushed with stored, pre-treated pollen at 2100 hrs that had been collected from Plant I on the morning of 05.07.21.
07.07.21 – Plant III, pollen extruded starting 2300 hrs, no temperatures taken. Spadix tip stored in referigeration but later discarded.
09.07.21 – Plants I and II returned to greenhouse.
14.07.21 – Plant III returned to greenhouse.
16.07.21 – Plants I and II drop pollinated inflorescences.
06.08.21 (30 days post pollination) – Plant III visible evidence of a successful pollination as staminate portion of infl. withers, berries begin to swell and change color from white to pale green.


Shown above left, infructescence 30 days post-pollination. Most of the fertile staminate section of the spadix was removed the day after pollination. Right, infructescence shown 48 days post-pollination after the staminate section of the spadix had dried and disintegrated. Author’s plant and images.
24.08.21 (48 days post pollination) - Plant III, remnant staminate portion of the spadix disintegrates.
12.09.21 (67 days post pollination) – Plant III, additional signs the berries are swelling. Note: Healthy-looking infructescences on members of this Philodendron subsection as well as those in subsections Fibrosa, Velveta and Canniphyllum that persist after 60 days post-pollination will usually complete the berry ripening process (A. Dearden, pers. comm.; pers. obs.).
13.10.21 (98 days post-pollination) - Plant III, basal portion of the spathe split by 1530 hrs.
14.10.21-15.10.21 (99-100 days post pollination) – Plant III, spathe decayed overnight and disintegrated from the base in the morning of the following day.


Shown above right, ripe infructescence of Philodendron sp. nov. “Chocó Red” at 100 days post-pollination and immediately following spathe dehiscence. Note irregular shaped berries and color change from light green to white from the previous gallery. Right, freshly harvested infructescence early the next day with soft ripe berries beginning to separate. Author’s images.
16.10.21 (101 days post pollination) – Plant III, infructescence harvested with soft, fully developed berries.
16.10.21 - Plant III, berries cleaned with a fine stainless-steel sieve and running water. Pulp was carefully decanted, and berries rinsed with mineral free tap water until clean in a 32 oz/945 ml clear plastic cup.
18.10.21-20.10.21 – Plant III, seed color begins to change from white to green; embryos starting to become visible in translucent seed on 19.01.21; seed uniformly pale green on 20.10.21.

Clean, sterile seed of Philodendron sp. nov. “Chocó Red” shown at magnification five days post-harvest and immediately prior to sow. Several hundred viable seed are visible in this photograph. Author’s image.
21.10.21 – Plant III, dilute bleach-sterilized seed (5% concentration for 10 minutes, followed by multiple rinses with distilled water) sown on steam-sterilized NZ sphagnum. Container covered with food grade plastic film held in place with an elastic band.
Low power microscopic examination was made prior to seed sow to gauge viability. Green embryos visible in many.

Seed shown in previous photo sown on heat sterilized New Zealand long fiber sphagnum moss in a 6”/15 cm bulb pan and covered with plastic wrap to maintain a humid microenvironment and to facilitate observation of seedling development. Author’s image.
Past experience shows that very fresh, viable Philodendron sect. Philodendron seed germinates rapidly (<21 days) and at almost 100% success rates if kept warm and shaded.
EDIT November 07, 2021: First radicle emergence noted on 05.11.21, 20 days post harvest, on a control sample held back for observation. By 07.11.21, ca 50% of the control seed showed visible germination.
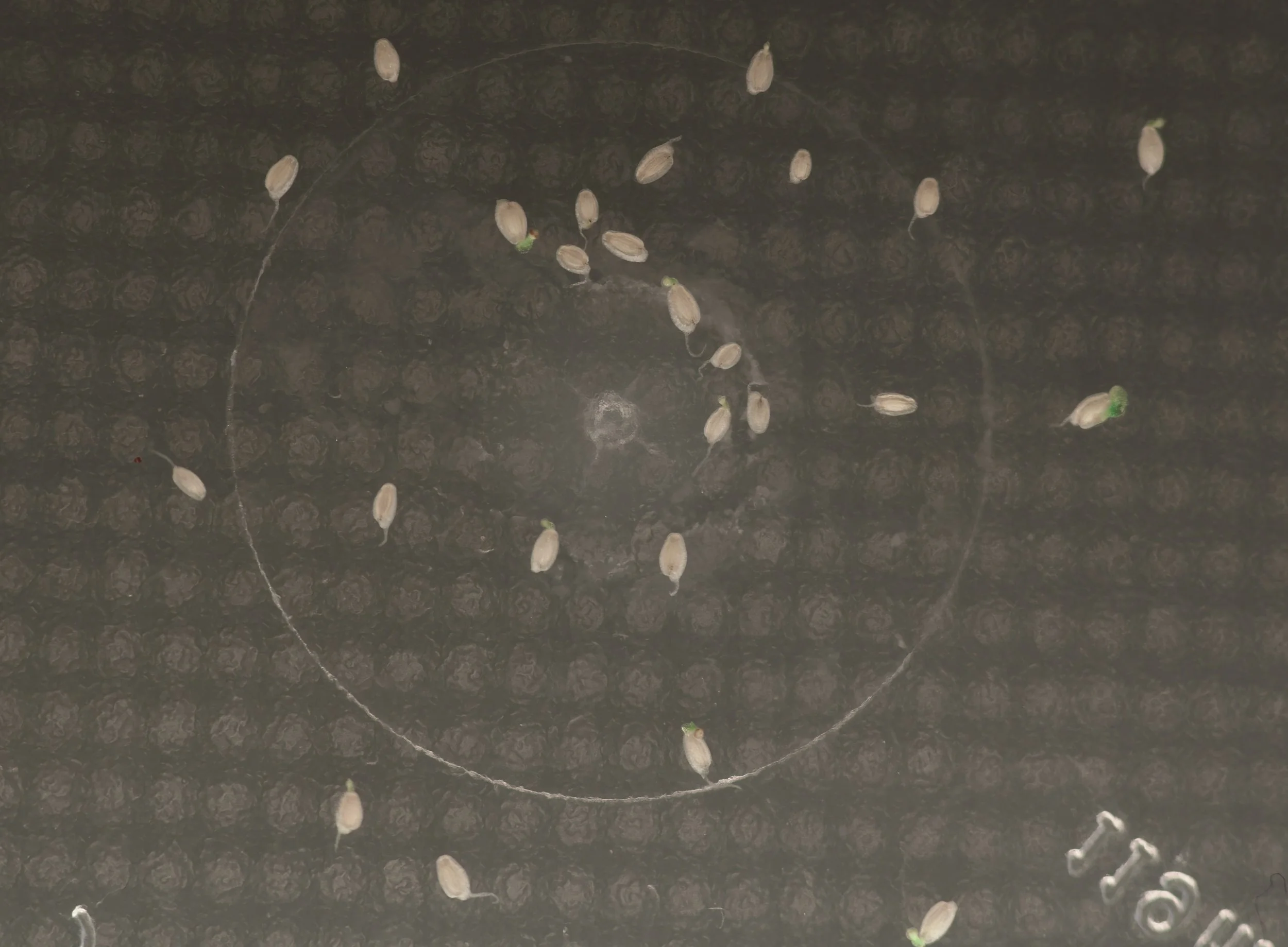
Germinating seeds of Philodendron sp. nov. “Chocó Red” control sample shown at magnification 22 days post harvest. Note seed color change from that shown above in freshly harvested seed. Author’s image.
EDIT November 21, 2021: Seedling leaves (ca. 1.50-2.50 mm diameter) visible throughout the compot, 31 days post seed sow.

Partially pollinated infructescence dimensions: 55 x 40 mm (wide), 70 mm in length overall including the incompletely pollinated basal portion of the spadix.
Berries: creamy white in color, irregularly shaped and 4 to 5 mm wide. Several seed per berry.
Seed description: ellipsoid-cuneate, ca. 1.0 mm x 0.5 mm. Freshly harvested seed color is white-translucent.
Acknowledgements
Thanks to Dr. Thomas Croat of the Missouri Botanical Garden and Dr. Ron Kaufmann of the University of San Diego for our spirited email exchanges from last fall and this spring regarding the likely identity of the enigmatic 19th century Philodendron triumphans. While we agreed to disagree–the point will soon be rendered moot in my opinion with the publication of their species diagnosis–the dialogues and examination of comparative plant material as well as alternative candidates for this classic name proved very interesting, at least for me.
Thanks also to Chris Hall and Arden Dearden of Queensland, Australia who shared very valuable information on their in-house Philodendron seed production protocols and proved to be excellent sounding boards as to how best to achieve pollination success with philodendrons that produce short-lived, resinous pollen. While our Philodendron breeding and seedling care techniques differ, I have no doubt that their input was critical to achieving the success reported here and am very grateful for this collaboration.
References Cited
Croat, T. B. 1997. A Revision of Philodendron subgenus Philodendron (Araceae) for Mexico and Central America. Annals of the Missouri Botanical Garden. Vol. 84. No. 3. pp 311-704.
Gardener’s Chronicle. 1898 xxiii. Haymarket Publishing. London. Pg. 254.
Gibernau, M. 2002. Pollination ecology of Philodendron squamiferum (Araceae), Canadian Journal of Botany. 80 (3): 316-320.
Graf, A. B. 1980. Exotica: Pictorial Cyclopedia of Exotic Plants from Tropical and Near-tropic Regions. 10th Edition. Roehrs Company Inc. 1833 pg.
Griffith Jr., L. P. 2001. Tropical Foliage Plants: A Grower’s Guide. Ball Publishing. 324 pg.
Hannon, D. P. 2014. Species Profile: Philodendron patriciae Croat (Araceae). Aroideana Vol. 37, No. 1: 105-110.
Henny, R. J., D. J. Norman and J. Chen. 2004. Progress in Ornamental Aroid Breeding Research. Annals of the Missouri Botanical Gardenm Vol. 91, No. 3. Pp. 467-472.
Maia, A. C. D., C. Schlindwein, D. Maria Almeida Ferraz Navarro and M. Gibernau. 2010. Pollination of Philodendron acutatum (Araceae) in the Atlantic Forest of Northeastern Brazil: A Single Scarab Beetle Species Guarantees High Fruit Set. International Journal of Plant Sciences. Vol. 171, No. 7. Sept. 2010. Pp. 740-748.
Miller L. M. and R.D. Newton. 2006. Hand Pollination and Seed Propagation of Philodendron bipinnatidum, Syn. P. selloum. Proc. Fla. State Hort. Soc. 119: 425-428. (Now Thaumatophylum bipinnatifidum).
Sakuragui, C M., L. S. Braucks Calazans, L. L. de Oliveira, E. B. de Morais, A. M. Benko-Iseppon, S. Vasconcelos, C. E. Guerra Schrago and S. J. Mayo. 2018. Recognition of the genus ThaumatophyllumSchott–formerly Philodendron subg. Meconostigma (Araceae)–based on molecular and morphological evidence. Phytokeys 98: 51-71.
Vannini, J. P. 2020. The Philo Philes – Part I. Satin & Velvet. Exotica Esoterica. Link: https://www.exoticaesoterica.com/exotic-plant-blog/2020/9/7/the-philo-files-part-i

A healthy seedling crop of artificially propagated Philodendron melanochrysum at 10 months post seed sow in a 6”/15 cm bulb pan. Author’s plants and images.
All content ©Exotica Esoterica LLC® and ©Jay Vannini 2021.
Follow us on:




